
百年の名作を、誰も読まなかった角度から
読まずに、深く知る。
読みたくなって、帰る。
世界の名作を、あらすじではなく「発見」から読み解く批評ライブラリ。原文を実際に数え、異分野の体系を当て、定説が見落としてきた構造を取り出します。
102おすすめ作品
1500収録作品

作品から入る
読まれ続けてきた名作から、最初に触れたい作品を選んでいます。
 戦争と平和ナポレオン戦争に巻き込まれる人びとの恋愛、家族、迷いを通じて、歴史が一人の英雄だけで動くわけではないことを読む大長編。戦場の命令、偶然、民衆の日常が重なり、時代が動いていく。
戦争と平和ナポレオン戦争に巻き込まれる人びとの恋愛、家族、迷いを通じて、歴史が一人の英雄だけで動くわけではないことを読む大長編。戦場の命令、偶然、民衆の日常が重なり、時代が動いていく。 赤毛のアン手違いで孤児院から引き取られた、痩せた赤毛のおしゃべりな少女。働き手の男の子が欲しかった老兄妹のもとへ、間違って届いた「不良在庫」のはずの彼女が、想像力という一つの才能だけで、無愛想な家と村を作り変えていく。これは可愛い少女物語ではなく、何も持たない者が想像力で世界の意味を書き換える物語である。
赤毛のアン手違いで孤児院から引き取られた、痩せた赤毛のおしゃべりな少女。働き手の男の子が欲しかった老兄妹のもとへ、間違って届いた「不良在庫」のはずの彼女が、想像力という一つの才能だけで、無愛想な家と村を作り変えていく。これは可愛い少女物語ではなく、何も持たない者が想像力で世界の意味を書き換える物語である。- 白鯨片脚を奪われた船長エイハブが、一頭の白い鯨に「宇宙の悪意」を読み込み、会社の船と乗組員の命を丸ごと私的な復讐に注ぎ込んで沈んでいく。これは鯨の物語ではない。意味のないものに意味を読みすぎた人間が、組織ごと滅びる物語である。
 不思議の国のアリス少女が穴に落ち、薬で伸び縮みし、意味の通らない住人たちに振り回される——一見ただの愉快なナンセンスだが、作者は数学者だった。ここで壊れているのは現実ではなく「論理と言語のルール」であり、アリスの冒険は、規則が信用できなくなった世界を子どもがサバイバルする記録である。
不思議の国のアリス少女が穴に落ち、薬で伸び縮みし、意味の通らない住人たちに振り回される——一見ただの愉快なナンセンスだが、作者は数学者だった。ここで壊れているのは現実ではなく「論理と言語のルール」であり、アリスの冒険は、規則が信用できなくなった世界を子どもがサバイバルする記録である。 グレート・ギャツビー貧しい青年が、自分を締め出した富そのものを体現する女を取り戻すために、巨万の富を持つ別人に成りすます。だが手が届いた瞬間、夢は色あせる。これは恋愛小説ではない。手に入れた瞬間に価値を失う「夢」という構造の解剖であり、アメリカン・ドリームの検死報告書である。
グレート・ギャツビー貧しい青年が、自分を締め出した富そのものを体現する女を取り戻すために、巨万の富を持つ別人に成りすます。だが手が届いた瞬間、夢は色あせる。これは恋愛小説ではない。手に入れた瞬間に価値を失う「夢」という構造の解剖であり、アメリカン・ドリームの検死報告書である。 レ・ミゼラブルパン一切れを盗んだ罪で19年を奪われた男が、たった一度の「赦し」によって人生を作り変えようとする。それを「犯罪者は一生犯罪者だ」と信じる警官が執拗に追う。これは波乱万丈の物語の顔をした、法と赦し(恩寵)の正面衝突の記録である。
レ・ミゼラブルパン一切れを盗んだ罪で19年を奪われた男が、たった一度の「赦し」によって人生を作り変えようとする。それを「犯罪者は一生犯罪者だ」と信じる警官が執拗に追う。これは波乱万丈の物語の顔をした、法と赦し(恩寵)の正面衝突の記録である。 ロミオとジュリエット二つの名家の若い男女が出会い、結婚し、すれ違い、四日でともに死ぬ。だがこの悲劇を殺したのは家の憎しみでも運命でもない。一通の手紙が届かなかったこと——それだけである。世界一有名な恋愛悲劇は、その実、情報伝達の事故の物語だ。
ロミオとジュリエット二つの名家の若い男女が出会い、結婚し、すれ違い、四日でともに死ぬ。だがこの悲劇を殺したのは家の憎しみでも運命でもない。一通の手紙が届かなかったこと——それだけである。世界一有名な恋愛悲劇は、その実、情報伝達の事故の物語だ。 若きウェルテルの悩み婚約者のいる女性に恋した青年が、叶わぬ想いと社会への違和感を募らせ、ついに自ら命を絶つ——その全過程を本人の手紙だけで綴る。これは失恋の物語である以上に、「感情を何より優先する」という新しい生き方が生まれた瞬間の記録であり、刊行が現実の自殺者を生んだ、文学史上もっとも危険な本である。
若きウェルテルの悩み婚約者のいる女性に恋した青年が、叶わぬ想いと社会への違和感を募らせ、ついに自ら命を絶つ——その全過程を本人の手紙だけで綴る。これは失恋の物語である以上に、「感情を何より優先する」という新しい生き方が生まれた瞬間の記録であり、刊行が現実の自殺者を生んだ、文学史上もっとも危険な本である。 カラマーゾフの兄弟下劣な父親が殺され、三人の兄弟がそれぞれの形で疑われる——だがこれは犯人探しの物語ではない。「もし神がいないなら、すべては許されるのか」という一つの問いを、三兄弟の生き方を通して、あらゆる角度から検証する、人類最大級の思考実験である。
カラマーゾフの兄弟下劣な父親が殺され、三人の兄弟がそれぞれの形で疑われる——だがこれは犯人探しの物語ではない。「もし神がいないなら、すべては許されるのか」という一つの問いを、三兄弟の生き方を通して、あらゆる角度から検証する、人類最大級の思考実験である。 嵐が丘荒野の屋敷に拾われた素性の知れない少年が、義妹との結ばれぬ愛をこじらせ、二つの家を二世代にわたって破壊し尽くす——だがこれは恋愛小説ではない。「愛」と呼ばれる、所有と憎悪と自己同一が見分けのつかなくなった激情が、人間をどこまで壊すかを見届ける実験である。
嵐が丘荒野の屋敷に拾われた素性の知れない少年が、義妹との結ばれぬ愛をこじらせ、二つの家を二世代にわたって破壊し尽くす——だがこれは恋愛小説ではない。「愛」と呼ばれる、所有と憎悪と自己同一が見分けのつかなくなった激情が、人間をどこまで壊すかを見届ける実験である。 ツァラトゥストラはこう語った山で十年こもった賢者ツァラトゥストラが、人々に「神は死んだ」「超人を目指せ」と説いて回る——だがこれは無神論の宣言ではない。神という究極の価値が信じられなくなった世界で、人間はどうやって自分の足で意味を作り出して生きるか、という問いへの、詩のような応答である。
ツァラトゥストラはこう語った山で十年こもった賢者ツァラトゥストラが、人々に「神は死んだ」「超人を目指せ」と説いて回る——だがこれは無神論の宣言ではない。神という究極の価値が信じられなくなった世界で、人間はどうやって自分の足で意味を作り出して生きるか、という問いへの、詩のような応答である。 イリアス十年に及ぶトロイア戦争を描く——のではない。この壮大な叙事詩が描くのは、最強の英雄アキレウスが「怒り」に駆られてから、その怒りを手放すまでの、わずか数十日間だ。戦争そのものではなく、一人の人間の怒りと、その果てにたどり着く赦しを描いた、西洋文学最古の心の物語である。
イリアス十年に及ぶトロイア戦争を描く——のではない。この壮大な叙事詩が描くのは、最強の英雄アキレウスが「怒り」に駆られてから、その怒りを手放すまでの、わずか数十日間だ。戦争そのものではなく、一人の人間の怒りと、その果てにたどり着く赦しを描いた、西洋文学最古の心の物語である。 オデュッセイアトロイア戦争を終えた英雄が、故郷へ帰るだけの旅に十年を費やす——怪物、魔女、誘惑、難破。だがこの冒険譚の本当の主題は、英雄が「力で勝つ」のではなく「知恵と忍耐で生き延びて帰る」ことだ。戦って死ぬ英雄像から、耐えて帰る英雄像への転換を、西洋文学の最初期に成し遂げた作品である。
オデュッセイアトロイア戦争を終えた英雄が、故郷へ帰るだけの旅に十年を費やす——怪物、魔女、誘惑、難破。だがこの冒険譚の本当の主題は、英雄が「力で勝つ」のではなく「知恵と忍耐で生き延びて帰る」ことだ。戦って死ぬ英雄像から、耐えて帰る英雄像への転換を、西洋文学の最初期に成し遂げた作品である。 モンテ・クリスト伯前途有望な若き船乗りエドモン・ダンテスは、結婚式の日、無実の罪を着せられ、絶海の孤島の牢獄に投獄される。彼を陥れたのは、その地位や恋人を妬む者たちだった。十四年の獄中で、彼は老司祭から知識と、莫大な財宝の在処を授かる。脱獄し、財宝を手にした彼は、『モンテ・クリスト伯』と名を変え、自分を破滅させた者たちへの、緻密な復讐を開始する。だが、復讐を遂げた果てに彼が見出したのは——。壮大な復讐の物語を通して、正義と復讐、そして赦しの意味を問う、不朽の大ロマンである。
モンテ・クリスト伯前途有望な若き船乗りエドモン・ダンテスは、結婚式の日、無実の罪を着せられ、絶海の孤島の牢獄に投獄される。彼を陥れたのは、その地位や恋人を妬む者たちだった。十四年の獄中で、彼は老司祭から知識と、莫大な財宝の在処を授かる。脱獄し、財宝を手にした彼は、『モンテ・クリスト伯』と名を変え、自分を破滅させた者たちへの、緻密な復讐を開始する。だが、復讐を遂げた果てに彼が見出したのは——。壮大な復讐の物語を通して、正義と復讐、そして赦しの意味を問う、不朽の大ロマンである。 罪と罰貧しい元学生が「選ばれた非凡人は法を踏み越えてよい」という理論を証明するため、老女を斧で殺す。だが完全犯罪は成立しても、彼の身体と心が理論を拒絶しはじめる。これは犯罪と捜査の物語ではない。頭で立てた理論を、人間の身体が受けつけなかった記録である。
罪と罰貧しい元学生が「選ばれた非凡人は法を踏み越えてよい」という理論を証明するため、老女を斧で殺す。だが完全犯罪は成立しても、彼の身体と心が理論を拒絶しはじめる。これは犯罪と捜査の物語ではない。頭で立てた理論を、人間の身体が受けつけなかった記録である。 変身ある朝、目を覚ますと巨大な虫になっていた——だがこの小説の本当の恐怖は、虫になったことではない。虫になった男が真っ先に心配するのが「会社に遅刻する」ことであり、稼げなくなった瞬間に家族から厄介者として処理されていくことだ。これは変身の物語ではなく、人間が経済的機能に還元される過程の記録である。
変身ある朝、目を覚ますと巨大な虫になっていた——だがこの小説の本当の恐怖は、虫になったことではない。虫になった男が真っ先に心配するのが「会社に遅刻する」ことであり、稼げなくなった瞬間に家族から厄介者として処理されていくことだ。これは変身の物語ではなく、人間が経済的機能に還元される過程の記録である。 ジェーン・エア美しくも裕福でもない孤児の少女が、家庭教師という最も無力な立場にありながら、決して自分の尊厳を売らない——たとえ愛のためであっても。これは恋愛小説の顔をした、一人の女性が「私はあなたと対等の人間だ」と宣言する、自尊心の物語である。
ジェーン・エア美しくも裕福でもない孤児の少女が、家庭教師という最も無力な立場にありながら、決して自分の尊厳を売らない——たとえ愛のためであっても。これは恋愛小説の顔をした、一人の女性が「私はあなたと対等の人間だ」と宣言する、自尊心の物語である。 ドリアン・グレイの肖像美しい青年ドリアン・グレイは、自分の肖像画に「自分の代わりに歳を取り、罪を背負ってほしい」と願う。願いは叶い、彼自身は若く美しいまま、肖像画だけが醜く老い、罪に歪んでいく。外見は永遠に美しく、しかし魂は腐っていく——美と快楽だけを追い、良心を捨てた人間の行きつく先を描いた、唯美主義の寓話である。
ドリアン・グレイの肖像美しい青年ドリアン・グレイは、自分の肖像画に「自分の代わりに歳を取り、罪を背負ってほしい」と願う。願いは叶い、彼自身は若く美しいまま、肖像画だけが醜く老い、罪に歪んでいく。外見は永遠に美しく、しかし魂は腐っていく——美と快楽だけを追い、良心を捨てた人間の行きつく先を描いた、唯美主義の寓話である。 善悪の彼岸私たちが「善い」「悪い」と呼ぶものは、永遠の真理なのか、それとも誰かが作り、私たちが受け継いだ道具にすぎないのか——ニーチェは、道徳そのものを疑いの対象に置く。善悪を当たり前の前提として使うのをやめ、その「向こう側(彼岸)」に立って、道徳がどこから来たのかを問い直す、危険で刺激的な思考の書である。
善悪の彼岸私たちが「善い」「悪い」と呼ぶものは、永遠の真理なのか、それとも誰かが作り、私たちが受け継いだ道具にすぎないのか——ニーチェは、道徳そのものを疑いの対象に置く。善悪を当たり前の前提として使うのをやめ、その「向こう側(彼岸)」に立って、道徳がどこから来たのかを問い直す、危険で刺激的な思考の書である。 ハックルベリー・フィンの冒険家出した少年と、逃げた奴隷が、いかだでミシシッピ川を下る——少年は「奴隷の逃亡を助けるのは罪だ」と教えられて育った。だが土壇場で彼は「よし、それなら地獄へ行こう」と決意し、友を助ける。社会が教える『正しさ』と、心が知る『正しさ』が真っ向からぶつかる、アメリカ文学の良心の物語である。
ハックルベリー・フィンの冒険家出した少年と、逃げた奴隷が、いかだでミシシッピ川を下る——少年は「奴隷の逃亡を助けるのは罪だ」と教えられて育った。だが土壇場で彼は「よし、それなら地獄へ行こう」と決意し、友を助ける。社会が教える『正しさ』と、心が知る『正しさ』が真っ向からぶつかる、アメリカ文学の良心の物語である。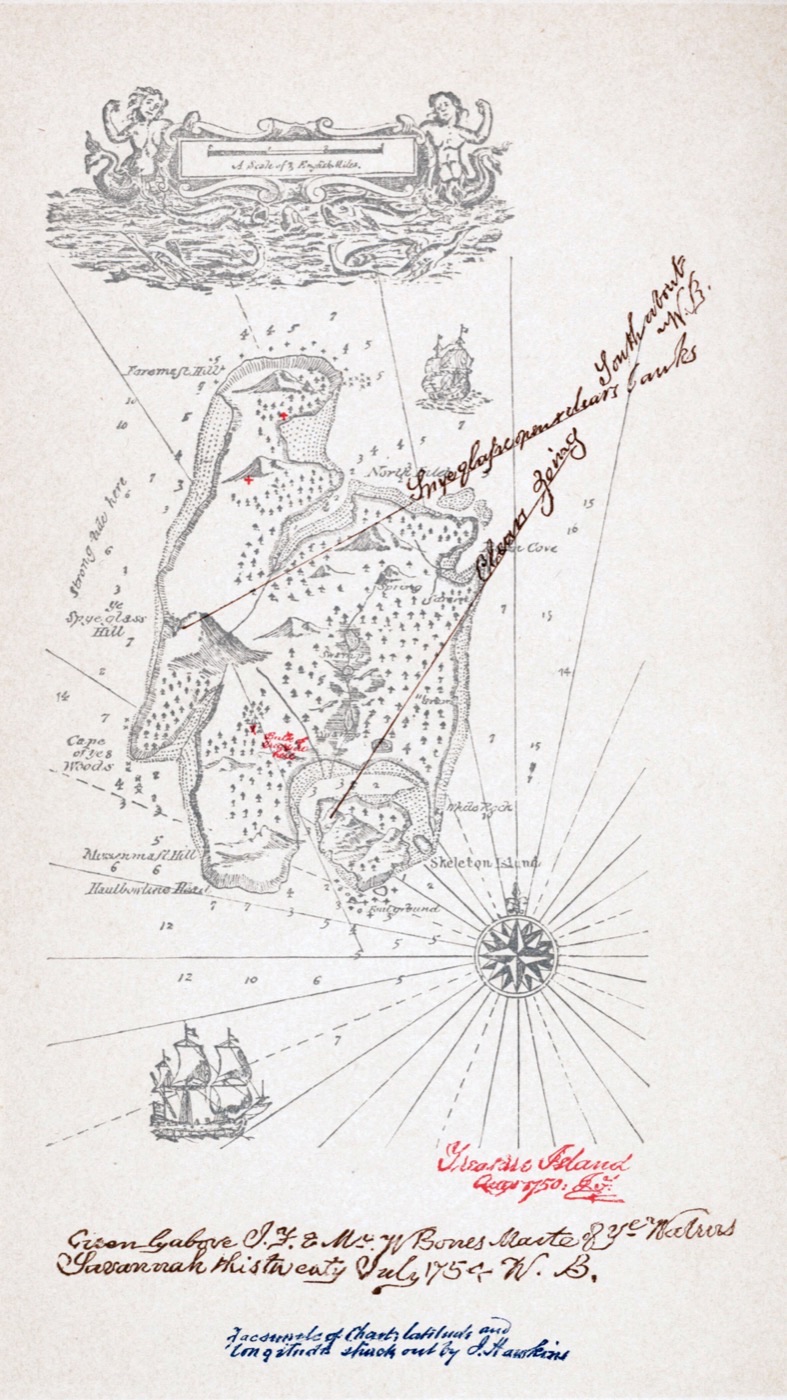 宝島宿屋の少年ジム・ホーキンズは、死んだ船乗りの荷物から、一枚の宝の地図を手に入れる。それは、伝説の海賊フリント船長が遺した、財宝の隠し場所を示すものだった。宝を求めて船出したジムだが、乗組員の多くは、料理人に化けた海賊たちで、その頭目が、片足の男ロング・ジョン・シルバー——魅力的で、優しく、しかし冷酷な、善とも悪ともつかぬ男だった。少年の目を通して、人間の善悪の曖昧さと、冒険の高揚を描いた物語である。
宝島宿屋の少年ジム・ホーキンズは、死んだ船乗りの荷物から、一枚の宝の地図を手に入れる。それは、伝説の海賊フリント船長が遺した、財宝の隠し場所を示すものだった。宝を求めて船出したジムだが、乗組員の多くは、料理人に化けた海賊たちで、その頭目が、片足の男ロング・ジョン・シルバー——魅力的で、優しく、しかし冷酷な、善とも悪ともつかぬ男だった。少年の目を通して、人間の善悪の曖昧さと、冒険の高揚を描いた物語である。 高慢と偏見機知に富み、誇り高い次女エリザベスと、裕福だが高慢に見える紳士ダーシー。二人は、最初の出会いで互いに反発しあう。エリザベスはダーシーの『高慢』を嫌い、ダーシーはエリザベスを見下す。だが、誤解と偏見の積み重ねの奥に、二人は、しだいに相手の本当の姿を見出していく。第一印象の『高慢』と『偏見』が、いかにして解け、本物の理解と愛へと変わっていくか——結婚をめぐる人間模様を通して、人が人を正しく知ることの難しさと喜びを描いた、恋愛小説の傑作である。
高慢と偏見機知に富み、誇り高い次女エリザベスと、裕福だが高慢に見える紳士ダーシー。二人は、最初の出会いで互いに反発しあう。エリザベスはダーシーの『高慢』を嫌い、ダーシーはエリザベスを見下す。だが、誤解と偏見の積み重ねの奥に、二人は、しだいに相手の本当の姿を見出していく。第一印象の『高慢』と『偏見』が、いかにして解け、本物の理解と愛へと変わっていくか——結婚をめぐる人間模様を通して、人が人を正しく知ることの難しさと喜びを描いた、恋愛小説の傑作である。 君主論君主は、いかにして権力を獲得し、維持すべきか——マキャヴェッリは、この問いに、道徳や理想を一切排して、冷徹な現実だけを見据えて答えた。『愛されるより恐れられよ』『必要とあらば、君主は約束を破り、残酷にもなれ』。人間は本来、利己的で移ろいやすいものだという前提から、彼は、きれいごとではない、生き残るための統治の技術を説く。善悪を超えて、政治の現実を直視したこの書は、発表当時から非難を浴びながらも、近代政治学の出発点となった、危険にして不朽の古典である。
君主論君主は、いかにして権力を獲得し、維持すべきか——マキャヴェッリは、この問いに、道徳や理想を一切排して、冷徹な現実だけを見据えて答えた。『愛されるより恐れられよ』『必要とあらば、君主は約束を破り、残酷にもなれ』。人間は本来、利己的で移ろいやすいものだという前提から、彼は、きれいごとではない、生き残るための統治の技術を説く。善悪を超えて、政治の現実を直視したこの書は、発表当時から非難を浴びながらも、近代政治学の出発点となった、危険にして不朽の古典である。 フランケンシュタイン若き科学者ヴィクター・フランケンシュタインは、生命の神秘を解き明かし、自らの手で生命を創り出すことに成功する。だが、つぎはぎの死体から生まれたその『怪物』のおぞましい姿に、彼は恐怖し、自分が創った存在を、見捨てて逃げ出す。誰からも愛されず、醜さゆえに憎まれ、孤独に苦しんだ怪物は、やがて、自分を捨てた創造主への、激しい復讐へと向かう——。科学者の傲慢、創造主の責任、そして『怪物』とは本当は誰なのかを問う、ゴシック小説にして世界最初のSFと呼ばれる、不朽の傑作である。
フランケンシュタイン若き科学者ヴィクター・フランケンシュタインは、生命の神秘を解き明かし、自らの手で生命を創り出すことに成功する。だが、つぎはぎの死体から生まれたその『怪物』のおぞましい姿に、彼は恐怖し、自分が創った存在を、見捨てて逃げ出す。誰からも愛されず、醜さゆえに憎まれ、孤独に苦しんだ怪物は、やがて、自分を捨てた創造主への、激しい復讐へと向かう——。科学者の傲慢、創造主の責任、そして『怪物』とは本当は誰なのかを問う、ゴシック小説にして世界最初のSFと呼ばれる、不朽の傑作である。 ドラキュライギリスの青年弁護士ジョナサン・ハーカーは、不動産取引のため、トランシルヴァニアの古城に住むドラキュラ伯爵を訪ねる。だがその城で、彼は伯爵が、夜に生き、人の血を吸う、不死の吸血鬼であることを知る。やがて伯爵はイギリスへと渡り、人々を餌食にしていく。これに立ち向かうのは、ヴァン・ヘルシング教授を中心とする人々だ。書簡や日記を積み重ねる形式で語られる、近代文明と古い闇との対決——人間の理性と科学が、いかにして説明のつかない『恐怖』と戦うかを描いた、吸血鬼物語の決定版である。
ドラキュライギリスの青年弁護士ジョナサン・ハーカーは、不動産取引のため、トランシルヴァニアの古城に住むドラキュラ伯爵を訪ねる。だがその城で、彼は伯爵が、夜に生き、人の血を吸う、不死の吸血鬼であることを知る。やがて伯爵はイギリスへと渡り、人々を餌食にしていく。これに立ち向かうのは、ヴァン・ヘルシング教授を中心とする人々だ。書簡や日記を積み重ねる形式で語られる、近代文明と古い闇との対決——人間の理性と科学が、いかにして説明のつかない『恐怖』と戦うかを描いた、吸血鬼物語の決定版である。 若草物語南北戦争のさなか、父を戦地に送り出したマーチ家の四姉妹——気が強く作家志望のジョー、優しく家庭的なメグ、内気で病弱なベス、美しくわがままなエイミー。決して豊かではない暮らしの中で、四人はそれぞれの欠点と向き合い、喜びと悲しみを分かち合いながら、少女から女性へと成長していく。特別な事件ではなく、家庭の日常そのものを通して、人が成長するとは何かを描いた、温かな物語である。
若草物語南北戦争のさなか、父を戦地に送り出したマーチ家の四姉妹——気が強く作家志望のジョー、優しく家庭的なメグ、内気で病弱なベス、美しくわがままなエイミー。決して豊かではない暮らしの中で、四人はそれぞれの欠点と向き合い、喜びと悲しみを分かち合いながら、少女から女性へと成長していく。特別な事件ではなく、家庭の日常そのものを通して、人が成長するとは何かを描いた、温かな物語である。 オズの魔法使い竜巻に家ごと吹き飛ばされ、不思議の国オズに迷い込んだ少女ドロシー。故郷カンザスへ帰る方法を求めて、彼女は『エメラルドの都』の大魔法使いオズのもとを目指す。道中で出会うのは、脳が欲しいかかし、心が欲しいブリキの木こり、勇気が欲しい臆病なライオン。だが旅の果てに彼らが気づくのは——求めていたものは、実は最初から自分の中にあった、という真実だった。自分の価値は、すでに自分の中にある。誰の心にも響く、自己発見の物語である。
オズの魔法使い竜巻に家ごと吹き飛ばされ、不思議の国オズに迷い込んだ少女ドロシー。故郷カンザスへ帰る方法を求めて、彼女は『エメラルドの都』の大魔法使いオズのもとを目指す。道中で出会うのは、脳が欲しいかかし、心が欲しいブリキの木こり、勇気が欲しい臆病なライオン。だが旅の果てに彼らが気づくのは——求めていたものは、実は最初から自分の中にあった、という真実だった。自分の価値は、すでに自分の中にある。誰の心にも響く、自己発見の物語である。 穏健なる提案穏健なる提案は、入口の違和感から読後に残る問いへ進むなかで、王権が人間の判断をどう変えるかを読む児童文学。
穏健なる提案穏健なる提案は、入口の違和感から読後に残る問いへ進むなかで、王権が人間の判断をどう変えるかを読む児童文学。 ハリエット・ジェイコブズ自伝ハリエット・ジェイコブズ自伝は、時代の入口から記憶としての結末へ進むなかで、選択が人間の判断をどう変えるかを読む伝記。
ハリエット・ジェイコブズ自伝ハリエット・ジェイコブズ自伝は、時代の入口から記憶としての結末へ進むなかで、選択が人間の判断をどう変えるかを読む伝記。 ドン・キホーテ騎士道物語を読みふけるあまり、現実と物語の区別がつかなくなった老郷士が、自らを遍歴の騎士『ドン・キホーテ』と名乗り、痩せ馬にまたがって、世の不正を正す冒険の旅に出る。風車を巨人と思い込んで突進し、宿屋を城と信じ込む——彼の『狂気』は、行く先々で滑稽な騒動を巻き起こす。だが、現実を理想で塗り替え、誰も信じない正義を一人本気で追い求めるその姿は、滑稽でありながら、いつしか胸を打つ気高さを帯びていく。狂気と理想、笑いと哀しみが分かちがたく溶け合う、近代小説の出発点である。
ドン・キホーテ騎士道物語を読みふけるあまり、現実と物語の区別がつかなくなった老郷士が、自らを遍歴の騎士『ドン・キホーテ』と名乗り、痩せ馬にまたがって、世の不正を正す冒険の旅に出る。風車を巨人と思い込んで突進し、宿屋を城と信じ込む——彼の『狂気』は、行く先々で滑稽な騒動を巻き起こす。だが、現実を理想で塗り替え、誰も信じない正義を一人本気で追い求めるその姿は、滑稽でありながら、いつしか胸を打つ気高さを帯びていく。狂気と理想、笑いと哀しみが分かちがたく溶け合う、近代小説の出発点である。 ジキル博士とハイド氏高名で人格者の医師ジキル博士には、ある秘密があった。彼は、自ら調合した薬によって、自分の中の『悪』だけを切り離し、醜く邪悪な別人格——ハイド氏——に変身できるようになっていたのだ。ハイドの姿になれば、ジキルは、世間体や良心に縛られず、欲望のままに振る舞える。だが、変身を繰り返すうち、ハイドはしだいに力を増し、ついにはジキルの意志を超えて、彼を乗っ取っていく。一人の人間の中に共存する善と悪、そして、抑圧された悪を解き放つことの恐ろしさを描いた、二重人格物語の原典である。
ジキル博士とハイド氏高名で人格者の医師ジキル博士には、ある秘密があった。彼は、自ら調合した薬によって、自分の中の『悪』だけを切り離し、醜く邪悪な別人格——ハイド氏——に変身できるようになっていたのだ。ハイドの姿になれば、ジキルは、世間体や良心に縛られず、欲望のままに振る舞える。だが、変身を繰り返すうち、ハイドはしだいに力を増し、ついにはジキルの意志を超えて、彼を乗っ取っていく。一人の人間の中に共存する善と悪、そして、抑圧された悪を解き放つことの恐ろしさを描いた、二重人格物語の原典である。 トム・ソーヤーの冒険トム・ソーヤーの冒険は、入口の違和感から読後に残る問いへ進むなかで、船が人間の判断をどう変えるかを読む児童文学。
トム・ソーヤーの冒険トム・ソーヤーの冒険は、入口の違和感から読後に残る問いへ進むなかで、船が人間の判断をどう変えるかを読む児童文学。 ユードルフォの秘密ユードルフォの秘密は、入口の違和感から読後に残る問いへ進むなかで、戦争が人間の判断をどう変えるかを読む小説。
ユードルフォの秘密ユードルフォの秘密は、入口の違和感から読後に残る問いへ進むなかで、戦争が人間の判断をどう変えるかを読む小説。 マンクマンクは、入口の違和感から読後に残る問いへ進むなかで、選択が人間の判断をどう変えるかを読む小説。
マンクマンクは、入口の違和感から読後に残る問いへ進むなかで、選択が人間の判断をどう変えるかを読む小説。 ユリシーズユリシーズは、入口の違和感から読後に残る問いへ進むなかで、船が人間の判断をどう変えるかを読む小説。
ユリシーズユリシーズは、入口の違和感から読後に残る問いへ進むなかで、船が人間の判断をどう変えるかを読む小説。 ピーター・パンとウェンディピーター・パンとウェンディは、入口の違和感から読後に残る問いへ進むなかで、旅が人間の判断をどう変えるかを読む児童文学。
ピーター・パンとウェンディピーター・パンとウェンディは、入口の違和感から読後に残る問いへ進むなかで、旅が人間の判断をどう変えるかを読む児童文学。 地底旅行地底旅行は、入口の違和感から読後に残る問いへ進むなかで、旅が人間の判断をどう変えるかを読む小説。
地底旅行地底旅行は、入口の違和感から読後に残る問いへ進むなかで、旅が人間の判断をどう変えるかを読む小説。 マノン・レスコーマノン・レスコーは、入口の違和感から読後に残る問いへ進むなかで、船が人間の判断をどう変えるかを読む小説。
マノン・レスコーマノン・レスコーは、入口の違和感から読後に残る問いへ進むなかで、船が人間の判断をどう変えるかを読む小説。 ガリヴァー旅行記ガリヴァー旅行記は、入口の違和感から読後に残る問いへ進むなかで、旅が人間の判断をどう変えるかを読む小説。
ガリヴァー旅行記ガリヴァー旅行記は、入口の違和感から読後に残る問いへ進むなかで、旅が人間の判断をどう変えるかを読む小説。 チャイルド・ハロルドの巡礼チャイルド・ハロルドの巡礼は、声の立ち上がりから読後の響きへ進むなかで、旅が人間の判断をどう変えるかを読む詩・詩集。
チャイルド・ハロルドの巡礼チャイルド・ハロルドの巡礼は、声の立ち上がりから読後の響きへ進むなかで、旅が人間の判断をどう変えるかを読む詩・詩集。 シャーロック・ホームズの冒険シャーロック・ホームズの冒険は、入口の違和感から読後に残る問いへ進むなかで、謎が人間の判断をどう変えるかを読む小説。
シャーロック・ホームズの冒険シャーロック・ホームズの冒険は、入口の違和感から読後に残る問いへ進むなかで、謎が人間の判断をどう変えるかを読む小説。 カルメンカルメンは、入口の違和感から読後に残る問いへ進むなかで、謎が人間の判断をどう変えるかを読む小説。
カルメンカルメンは、入口の違和感から読後に残る問いへ進むなかで、謎が人間の判断をどう変えるかを読む小説。 緋色の研究緋色の研究は、入口の違和感から読後に残る問いへ進むなかで、謎が人間の判断をどう変えるかを読む小説。
緋色の研究緋色の研究は、入口の違和感から読後に残る問いへ進むなかで、謎が人間の判断をどう変えるかを読む小説。 湖上の美人 (詩集)湖上の美人 (詩集)は、声の立ち上がりから読後の響きへ進むなかで、旅が人間の判断をどう変えるかを読む詩・詩集。
湖上の美人 (詩集)湖上の美人 (詩集)は、声の立ち上がりから読後の響きへ進むなかで、旅が人間の判断をどう変えるかを読む詩・詩集。 木曜日の男木曜日の男は、入口の違和感から読後に残る問いへ進むなかで、謎が人間の判断をどう変えるかを読む小説。
木曜日の男木曜日の男は、入口の違和感から読後に残る問いへ進むなかで、謎が人間の判断をどう変えるかを読む小説。 海賊海賊は、入口の違和感から読後に残る問いへ進むなかで、船が人間の判断をどう変えるかを読む小説。
海賊海賊は、入口の違和感から読後に残る問いへ進むなかで、船が人間の判断をどう変えるかを読む小説。 ねじの回転ねじの回転は、入口の違和感から読後に残る問いへ進むなかで、謎が人間の判断をどう変えるかを読む児童文学。
ねじの回転ねじの回転は、入口の違和感から読後に残る問いへ進むなかで、謎が人間の判断をどう変えるかを読む児童文学。 四つの署名四つの署名は、入口の違和感から読後に残る問いへ進むなかで、謎が人間の判断をどう変えるかを読む小説。
四つの署名四つの署名は、入口の違和感から読後に残る問いへ進むなかで、謎が人間の判断をどう変えるかを読む小説。
ニュースを古典で読む
ニュースの要約ではなく、その日の出来事から古典の構造と今後の示唆へ進むシリーズです。
- なぜ、値段が少し下がっても暮らしの不安は消えないのか2026年6月26日、AP通信は、米連邦準備制度理事会が重視する物価指標が5月に前年同月比4.1%となり、3年ぶりの高水準になったと報じた。ガソリン価格はその後やや落ち着いても、暮らしの不安はすぐには軽くならない。ここで読むべきなのは、物価が上がったという一言だけではない。社会はしばしば、ある品目の値下がりより前に、家計の側で先に削られた安心の方を元に戻しにくいという構造である。国富論、クリスマス・キャロル、レ・ミゼラブルを並べると、このニュースは統計の速報ではなく、価格と生活のあいだにある時間差を読む入口になる。
- 逃れた人の居場所はなぜ、仮のままにされるのか2026年6月25日、AP通信は、米連邦最高裁がハイチ人とシリア人向けの一時保護資格の打ち切りを認め、少なくとも35万人のハイチ人と約6000人のシリア人が送還の危険に直面すると報じた。ここで読むべきなのは、移民政策がまた厳しくなったという一言ではない。国家はしばしば、逃れてきた人に屋根と仕事を与えても、その居場所を正式な帰属へ変えず、いつでも剥がせる仮住まいのまま保ちたがるという構造である。オデュッセイア、レ・ミゼラブル、リヴァイアサンを並べると、このニュースは法廷の速報ではなく、人が家を持つことと国家に認められることがなぜ別問題のまま残り続けるのかを読む入口になる。
- 住まいの法案はなぜ、忠誠の試験に変わるのか2026年6月24日、AP通信は、トランプ大統領が超党派の住宅法案の署名式を直前で取りやめ、証明書類による市民権確認を求める投票法案の前進を条件にしたと報じた。ここで読むべきなのは、政争がまた一つ荒れたということだけではない。住まいの供給や信用へのアクセスといった生活の土台が、しばしば権力の忠誠試験に変わるという構造である。ハード・タイムズ、君主論、クリスマス・キャロルを並べると、住宅ニュースは政策の遅延ではなく、政治が人の暮らしをどこで交渉材料に変えるのかを読む入口になる。
- 通れるだけで平和にならないのはなぜか2026年6月23日、AP通信は、米国とイランの暫定合意後にホルムズ海峡の船舶通行は持ち直しつつある一方、海峡を誰が管理し、将来通行料を課すのかが次の交渉の火種になっていると報じた。ここで読むべきなのは、船が何隻通ったかだけではない。平和はしばしば停戦文書より後で試される。人と物が通る細い通路を、誰が守り、誰が値段をつけ、誰が恐怖を管理するのかという構造である。アナバシス、リヴァイアサン、国富論を並べると、海峡のニュースは中東の局地的な話ではなく、補給路と秩序と商業がどこで同じ綱を引き合うかを読む入口になる。
- 政権はなぜ、議席より先に空洞化するのか2026年6月22日、英国のキア・スターマー首相が辞任を表明し、後継が決まるまで暫定的に職にとどまるとAP通信が報じた。ここで読むべきなのは、一人の指導者の失敗談だけではない。政権はしばしば、選挙や解任という形式的な敗北より先に、味方がその人のもとで損を引き受けなくなった時点で空洞化する。社会契約論、アメリカのデモクラシー、虚栄の市を並べると、辞任のニュースは人事の速報ではなく、統治がどこで正当性を失うのかを読む入口になる。
- 言葉はなぜ、制度を静かに奪うのか2026年6月21日、チェコのプラハで、公共放送の財源を受信料から国家予算へ切り替える政府案への抗議が行われた。ここで読むべきなのは、放送局の予算論だけではない。制度はしばしば、閉鎖や検閲より先に、効率化や節約という穏やかな言葉で権力へ近づけられる。ガリヴァー旅行記、アメリカのデモクラシー、国家を並べると、メディアの独立とは理念ではなく、誰の機嫌から距離を取れる仕組みかという問題だと見えてくる。
- 大雨の前に、人はなぜ備えを後回しにするのか2026年6月21日、東北北部の梅雨入りが発表され、東北地方では大雨への警戒が呼びかけられた。ここで読むべきなのは、天気の解説だけではない。人間はなぜ危険を知っていても備えを遅らせるのかである。台風、海に働く人びと、白鯨を並べると、自然災害の問題は自然の強さだけでなく、人間が自分の判断を過信する構造として見えてくる。
- 合意はなぜ、次の争いの始まりになるのか2026年6月20日、米国のウィトコフ特使がスイスへ向かい、米国とイランの核問題をめぐる協議が近く始まる見通しだと報じられた。一方で、覚書の後も軍事衝突やホルムズ海峡をめぐる懸念は残り、収束は見通しにくい。ここで読むべきなのは、誰が何を言ったかだけではない。合意とは、争いの終点ではなく、各陣営が勝利、譲歩、裏切り、安全をどう解釈するかの始まりである。君主論、リヴァイアサン、戦争と平和を並べると、外交合意のニュースは、平和の宣言ではなく、権力と恐怖と世論が次にどう動くかを読む入口になる。
- 炎上はなぜ正義の顔をして広がるのかSNSの炎上は、単なる怒りの爆発ではない。人々は怒っているだけでなく、自分が正しい側にいることを確かめようとする。緋文字、虚栄の市、アメリカのデモクラシー、社会契約論を並べると、炎上とはデジタル時代だけの現象ではなく、共同体が誰かを罰することで自分の秩序を確認する古い構造だと見えてくる。
- 働いても豊かにならないのはなぜか働いているのに余裕がない。成果を出しても安心が増えない。これは現代だけの不満ではない。国富論、ハード・タイムズ、バートルビー、クリスマス・キャロルを並べると、働くことの問題は賃金だけではなく、人間が交換可能な部品として扱われるとき、生活の意味まで削られるという構造として見えてくる。
横断テーマから入る
- 偽装の系譜——ハムレットの狂気、葉蔵の道化、檸檬の悪漢一冊だけ読んでいては見えない発見がある。シェイクスピアの王子は狂気のふりをし、太宰の葉蔵は道化を演じ、梶井の「私」はなに食わぬ顔で爆弾犯のふりをする。300年離れた三つの「ふり」を一枚に並べると、偽装という生存技術の系譜図が浮かび上がり、三作の結末がなぜああなったのかが、一つの法則で説明できてしまう。
- 人はなぜ罪を語らずにいられないのか罪は、隠せば消えるものではない。ラスコーリニコフは理屈で自分を守ろうとし、カラマーゾフ家の人々は責任を押しつけ合い、先生は手紙でしか告白できず、イワン・イリイチは死の前で人生の嘘を見てしまう。四つの作品を並べると、罪とは法律違反だけではなく、自分の物語を最後まで引き受けられるかどうかの問題だと見えてくる。
- 怪物は本当は何を映しているのか怪物は、外から襲ってくるだけの存在ではない。フランケンシュタインの怪物は作った者の責任を映し、ドラキュラは共同体の恐怖を集め、ジキルとハイドは分けたはずの欲望の戻り方を見せ、ドリアン・グレイは美しさの裏にしまった腐敗を肖像へ押し込める。四つを並べると、怪物とは人間が自分の中に置けなかったものの姿だとわかる。
- 子どもの目には世界がどう見えるのか子どもが主人公の物語は、かわいい冒険では終わらない。アリスは大人の言葉の奇妙さを暴き、アンは想像力で孤独な現実を塗り替え、トム・ソーヤーは遊びで社会の規則をすり抜け、セーラは貧しさの中で尊厳を手放さない。四つを並べると、子どもの視点は未熟さではなく、大人の世界を相対化する批評の装置だと見えてくる。
- 恋はなぜ人の目を曇らせるのか恋愛小説は、好きになった二人の話だけではない。アンナは家庭と社交界の視線に追い詰められ、エマは物語の中の恋に現実を合わせようとし、エリザベスは偏見をほどき、ロミオとジュリエットは世界の敵意を恋の速度で突破しようとする。四つを並べると、恋とは相手を見る力であると同時に、世界を見誤る力でもあるとわかる。
- お金は人をどこまで変えるのかお金は、ただの交換手段ではない。国富論は社会を動かす見えない仕組みを語り、クリスマス・キャロルは金に閉じた心の回復を描き、大いなる遺産は財産が自己像を変える怖さを示し、互いの友は遺産とごみ山の周囲に人間関係が群がる様子を見せる。四つを読むと、お金は財布の中ではなく、人が自分をどう説明するかの中に入り込むものだとわかる。
- 政治を読むと暮らしはどう見えるのか政治思想の古典は、遠い制度論ではない。国家は正しさと教育を問い、君主論は権力の現実を冷たく見つめ、リヴァイアサンは秩序がなぜ必要かを考え、市民的不服従は従わないことの責任を示し、アメリカのデモクラシーは多数派の力と危うさを読む。五つを並べると、政治とは議会の外側にもあり、職場、学校、家庭の中で人がどう従い、どう逆らうかの問題だと見えてくる。
- 旅に出ると何がはがれ落ちるのか旅の物語は、遠くへ行く話に見えて、実は自分を支えていた前提がはがれる話である。白鯨では海が人間の執念を拡大し、八十日間世界一周では時間と計算が旅を支配し、地底旅行では地面の下に降りることで常識が崩れる。三つを並べると、旅とは景色を増やすことではなく、自分が何に縛られていたかを露出させる装置だとわかる。